动物界—裸子动物
实核生物域
动物界
裸子动物

裸子动物(学名:Gymnospermae)是指种子动物中,胚珠在一开放的胞子叶边沿或叶面的动物,胞子叶凡是会摆列成圆锥的外形;因为胞子叶其实不向内卷合包覆胚珠,所以也无子房的构造。
裸子动物共有5个门约14科88属超越一千种。裸子动物那个名称源自希腊语“gymnospermos”,意指“ *** 的种子”,因为裸子动物的胚珠外围没有子房壁庇护,故称做裸子动物。与之相对的是种子动物的另一次要类群被子动物,其胚珠则是在向内卷合的心皮(一个边沿相接的孢子叶)内遭到包被与庇护。
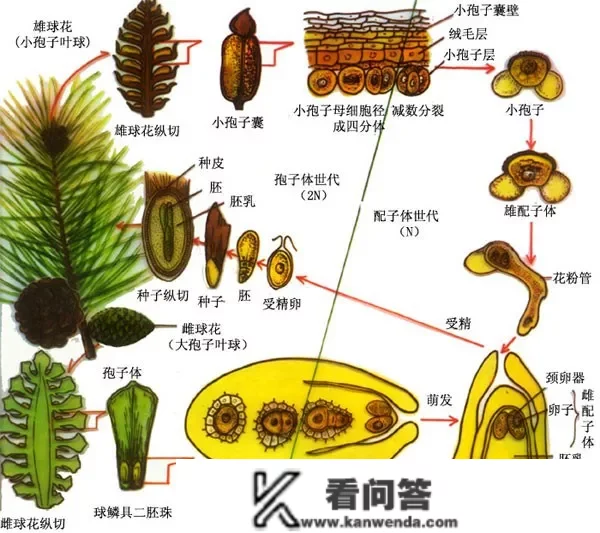
裸子动物会产生胞子,有开展成似花粉的“小胞子”和留在胚珠里的“大胞子”两种。当受精(大胞子和小胞子连系)之后,构成的胚芽便会和其他细胞构成胚珠,并开展成种子。
1.汗青
裸子动物发作开展的汗青悠久,最后的裸子动物呈现约在34500万年前至39500万年之间的古生代泥盆纪,历经古生代的石炭纪、二叠纪,中生代的三叠纪、侏罗纪、白垩纪,重生代的第三纪、第四纪。从裸子动物产生到21世纪,地量天气颠末屡次严重变革,裸子动物种系也随之屡次演变动替,老的品种相继灭绝,新的品种陆续演化出来,品种演替繁衍至今,现代的裸子动物有很多品种是从约250万年前至6500万年之间的重生代第三纪呈现的,又颠末第四纪冰川期间保留下来,繁衍至今。
较多学者认为裸子动物是由前裸子动物(Progymnospermae)和种子蕨(Pteridospermopsida,即苏铁蕨Cycadofilicopsida)演化而来。实蕨类(Filicopsida)与前裸子动物可能配合起源于裸蕨类(Psilophyta),而种子蕨类与其他裸子动物又平行起源于前裸子动物。科达类(Cordaitopsida)可能起源于前裸子动物,而本内苏铁类(Bennettitopsida,即拟苏铁类Cycadeoideopsida)可能起源于种子蕨类的皱叶羊齿类(Lyginopteridatae),苏铁类(Gycadopsida)与皱叶羊齿类亲缘关系亲近。银杏类(Ginkgopsida)与苛得狄类可能有配合的起源,它们可能是由前裸子动物的统一个分枝中演化出来,或一起头就相互独立演化成两个平行分枝。松杉类(Coniferopsida)与苛得狄类有着很近的亲缘关系。盖子动物(Chlamydospermopsida)是极特殊的类群,因缺乏古动物学材料,未见有阐明其起源与演化的报导。
2.形态特征
裸子动物门(Gymnospermae)是动物界的1门,既是颈卵器动物,又是种子动物,它们有胚珠(差别于蕨类动物门),但心皮不包成子房,且胚珠 *** ,胚乳(即雌性原叶体)在受精前已构成(差别于被子动物门)。
裸子动物为多年生木本动物,大多为单轴分枝的高峻乔木,少为灌木,稀为藤本;次生木量部几全由管胞构成,稀具导管。叶多为线形、针形或鳞形,稀为羽状全裂、扇形、阔叶形、带状或膜量鞘状。花单性,雌雄异株或同株;小胞子叶球(雄球花)具大都小胞子叶(雄蕊),小胞子叶具大都至2个小胞子囊(花药),小胞子(花粉)具气囊或船形具单沟,或球形外壁上具一 *** 状突起或具明显或不明显的萌生孔或无萌生孔,或橄榄形具多纵肋和凹沟,有时还具一远极沟,多为风媒传粉,花粉萌生后花粉管内有两个游动或不游动的 *** ;大胞子叶(珠鳞、珠托、珠领、套被)不构成封锁的子房,着生一至多枚 *** 的胚珠,大都丛生树干顶端或生于轴上构成大胞子叶球(雌球花);胚珠曲立或倒生,由胚囊、珠心和珠被构成,顶端有珠孔。种子 *** 于种鳞之上,或几被反常大胞子叶发育的假种皮所包,其胚由雌配子体的卵细胞受精而成,胚乳由雌配子体的其他部门发育而成,种皮由珠被发育而成;胚具两枚或多枚子叶。裸子动物的染色体基数少(x=8~20),形较大,在各属根本一致。
3.散布范畴
裸子动物广布于南北半球,尤以北半球更为普遍,从低海拔至高海拔、从低纬度至高纬度几乎都有散布。裸子动物的科、属、种数虽远比被子动物少,但笼盖面积却大致相等。在高纬度及高海拔天气温凉至冰冷的地域,几乎都是某些裸子动物构成的单纯林或构成的混交林。
裸子动物分为:苏铁科(Cycadaceae)、罗汉松科(Podocarpaceae)和南洋杉科(Araucariaceae),除其形式属(即苏铁属、罗汉松属、南洋杉属)的少数种散布于北半球热带及亚热带外,其他属种均产南半球;银杏原产中国,现普遍栽于北半球亚热带及温带地域;松科(Pinaceae)除松属(Pinus)的少数种散布于南半球外,其他属种均产北半球,此中油杉属、金钱松属、黄杉属、雪松属、银杉属、以及松属和铁杉属的部门品种散布于亚热带低山至中山地带,跟着纬度或海拔的升高,逐步被耐寒、喜温凉冷湿的少数松树、铁杉及落叶松属、云杉属和冷杉属树种所取代;杉科除单型属密叶杉属产澳大利亚外,其他属种均散布于北半球的亚热带地域;柏科散布于南北半球;三尖杉科散布于东亚南部及中南半岛北部;红豆杉科除澳洲红豆杉属产新喀里多尼亚外,其他属种均散布于北半球亚热带及温带;麻黄科散布于北半球温带及亚热带高山;买麻藤科散布于亚洲、非洲及南美洲的热带及亚热带地域;单型科百岁兰科散布于安哥拉及非洲热带东南部。
4.次要价值
裸子动物除少数类群(如买麻藤属及松科的一些属)外,均具有双黄酮类化合物,常见的有穗花杉双黄酮、西阿多黄素、银杏黄素、枯黄素、榧黄素等。黄酮类化合物则遍及存在,常见的有槲皮素、山奈酚和杨梅树皮素等。生物碱仅在三尖杉科、麻黄科和买麻藤科中存在,可供药用。苏铁科、红豆杉科和罗汉松科的部门属种可供欣赏,其他品种多为高峻乔木,其树干通曲,出材率高,材量较优良,供建筑、家具及工业用材,占世界木材供给量的50%以上;部门树种可割造松香和提取松节油;少数树种的种子可食;部门树种为速生造林树种或园林绿化树种;生于江河上游的为水源林;生于高山陡坡的可避免雨水冲刷。
我国的裸子动物多为林业运营上的重要用材树种,也是纤维、树脂、单宁等原料树种,少数品种的枝叶、花粉、种子、根皮等可供药用。

5.分类
在早期的分类里,裸子动物被认为是一个“天然”的群体。但是,一些化石的发现推测被子动物可能演化自一裸子动物的祖先,那将使得裸子动物构成一个并系群,若将所有灭绝的物种都考虑进来的话。现代的亲缘分收分类法只承受单系群的分类,可逃溯至一配合的祖先,且包罗着此一配合祖先的所有后代。因而,固然“裸子动物”一词仍然普遍地被利用来指非被子动物的其他种子动物,但之前一度被视为裸子动物的动物物种一般都被分至五个类群中,以让动物界内的门都有着不异的阶层。
考虑其他已灭绝的裸子动物,现存物种的分子种系发作学已和其关于开花动物是构成一单系群或并系群的形态类别相抵触。而还在争议上的还有,买麻藤门会是被子动物的旁收,亦或是其他已灭绝的裸子动物之旁收,或同源。
裸子动物包罗可能由差别道路进化所构成的若干平行的类群,故目前不再做为天然分类中的一个门,而将各纲递进为门,所以裸子动物包罗种子蕨门、苏铁门(本内苏门)、银杏门、松柏门。分类位置不定的买麻藤动物门有些学者也归入裸子动物。
种子蕨门始现于早石炭世,侏罗纪后绝灭。它兼具实蕨动物和典型裸子动物的特征,即具几与实蕨无异的大型羽状复叶,但角量层厚,并在叶上着生种子或雄性传粉器官,故名。动物体不大,主茎很少分枝,除部门为乔木或树蕨型外,大大都为较小的灌木或藤本。茎的剖解显示次生木量部薄而皮部厚,与苏铁动物类似,曾有苏铁蕨之称。归入此门的有良多目,皱羊齿目、髓木目、美籽目(芦茎羊齿目)、盾形种子目、兜状种子目。开通目,可能还有五柱木目及舌羊齿目亦列入种子蕨门。舌羊齿目为具生长轮的大树,具南美杉型裸子动物木材特征;单网状的舌形叶,生殖器官极为多样化,但仍着生于叶上。
苏铁门为已绝灭的类群,仅发现于晚三叠世至白垩纪。动物体与现代苏铁类似或更粗短。差别的是雌雄生殖器官着生于统一动物体内呈两性花状,表皮细胞及气孔器亦差别于苏铁类。苏铁动物门则自晚石炭世起就有过渡型化石发现,从到及表白苏铁目是由种子蕨动物演化而来的,化石证据也表白,在古生代、中生代苏铁动物曾有茎干纤细的类型。
银杏门现代仅有1属1种,为出名的活化石,但在中生代尤其是侏罗纪至早白垩世曾是广布北半球暖温带动物区系的重要成员,有些以至达南半球。确属本目标较早成员发现于早二叠世,近年来有人按照银杏动物门的雌性胚珠特点认为与种子蕨动物门的盾形种子目有关。本门自白垩纪末起,散布区及属种数量都锐减。
松柏门在早石炭世已有化石记录,属于已绝灭的科达目,该目呈乔木状,在晚石炭世晚期至早二叠世为热带动物区次要聚煤动物之一。伏脂杉目介于科达目与松柏目之间,生活于二叠纪至三叠纪,其重要的代表科属如勒巴杉科(Lebachiaceae),伏脂杉科(Voltziaceae)的生殖器官研究对由白垩世至今茂盛的松柏目标起源具有重要感化。现存的松柏目各科在中生代也都已发现切当的化石根据。
现行分类:
种子蕨门(Pteridospermatophyta)
松柏门(Pinophyta)
银杏门(Ginkgophyta)
苏铁门(Cycadophyta)
买麻藤门(Gnetophyta)
【更多出色文章,请存眷微信公家号“世界民族与文明汗青”】